 A finales del siglo XIX, tras el desarrollo de la teoría evolutiva basada en la selección natural, numerosos museos fueron creados tanto en Europa como en Norteamérica para exhibir al público los descubrimientos realizados por las cada vez más abundantes expediciones de recolección de fósiles, así como para fomentar su estudio científico. Es la época de paleontólogos tan reconocidos como Jean Albert Gaudry (1827-1908), Thomas Henry Huxley (1825-1895), Othniel Charles Marsh (1831-1899), Edward Drinker Cope (1840-1897), Henry Fairfield Osborn (1857-1935), Eugène Dubois (1858-1940) u Otto Jaekel (1863-1929), y de descubrimientos tan reputados como el esqueleto casi completo de Iguanodon de Bernissart, los especimenes de Archaeopteryx de Solnhofen, los innumerables dinosaurios del Norteamérica, la fauna miocena de Pikermi, los múltiples équidos del Terciario del Oeste de los Estados Unidos o los restos de “Pithecanthropus” en Java.
A finales del siglo XIX, tras el desarrollo de la teoría evolutiva basada en la selección natural, numerosos museos fueron creados tanto en Europa como en Norteamérica para exhibir al público los descubrimientos realizados por las cada vez más abundantes expediciones de recolección de fósiles, así como para fomentar su estudio científico. Es la época de paleontólogos tan reconocidos como Jean Albert Gaudry (1827-1908), Thomas Henry Huxley (1825-1895), Othniel Charles Marsh (1831-1899), Edward Drinker Cope (1840-1897), Henry Fairfield Osborn (1857-1935), Eugène Dubois (1858-1940) u Otto Jaekel (1863-1929), y de descubrimientos tan reputados como el esqueleto casi completo de Iguanodon de Bernissart, los especimenes de Archaeopteryx de Solnhofen, los innumerables dinosaurios del Norteamérica, la fauna miocena de Pikermi, los múltiples équidos del Terciario del Oeste de los Estados Unidos o los restos de “Pithecanthropus” en Java.
Sin embargo, y a pesar del relanzamiento de los estudios paleontológicos, una buena parte de la disciplina permaneció restringida el estudio morfológico e incluso las ideas evolutivas basadas en la selección natural no fueron plenamente aceptadas por la mayoría de los paleontólogos durante las primeras décadas del siglo XX (Bowler, 1990). Muchos trabajos se podrían adscribir a una especie de Neo-Lamarckismo basado en el concepto de recapitulación, según el cual los procesos ontogenéticos podían extenderse para dar lugar a procesos evolutivos. La existencia de evidencias de evolución paralela tuvo como consecuencia el desarrollo del concepto de ortogénesis, basado en la existencia de fuerzas internas a los organismos y que dirigían su evolución.
 Hubo de esperarse hasta la década de los 40, cuando George Gaylord Simpson (1902-1984) publicó "Tempo and Mode in Evolution" y se desarrolló plenamente la Síntesis Evolutiva Moderna, para que los paleontólogos abrazasen sin reparos los estudios evolutivos. A partir de ese momento proliferaron los trabajos que planteaban escenarios evolutivos basados en la selección natural y hubo un mayor interés por aspectos microevolutivos en poblaciones locales.
Hubo de esperarse hasta la década de los 40, cuando George Gaylord Simpson (1902-1984) publicó "Tempo and Mode in Evolution" y se desarrolló plenamente la Síntesis Evolutiva Moderna, para que los paleontólogos abrazasen sin reparos los estudios evolutivos. A partir de ese momento proliferaron los trabajos que planteaban escenarios evolutivos basados en la selección natural y hubo un mayor interés por aspectos microevolutivos en poblaciones locales.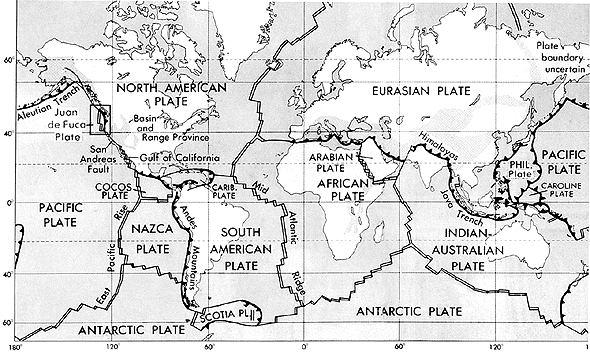 La aceptación en los años 60 del siglo XX y posterior desarrollo de la tectónica de placas supuso una nueva revolución teórica a la que la disciplina de la Paleontología tuvo que adaptarse, abandonando las propuestas de puentes terrestres e hundimientos continentales como maneras de explicar la distribución geográfica de los organismos actuales y pasados. Por otro lado, en el campo de la Biología se produjo un progresivo incremento del interés por los estudios ecológicos, en relación con la creciente percepción del impacto antrópico en los ecosistemas. Dentro de nuestra disciplina, el desarrollo de estas áreas de investigación tuvo como resultado un aumento de los estudios paleobiogeográficos y paleoecológicos (Valentine, 1990). Además, también comenzó por aquel entonces el estudio de las primeras formas de vida terrestre (Awramik, 1971) y se confirmó la preeminencia de la biota de Ediacara (Glaessner, 1959), lo cual propulsó los estudios de fósiles precámbricos. Esas son también las fechas de la reapertura de las excavaciones en Burgess Shale (Briggs et al., 1994; Conway Morris, 1998). Estos trabajos pusieron de manifiesto la complejidad de los eventos evolutivos que dieron lugar a los filos modernos.
La aceptación en los años 60 del siglo XX y posterior desarrollo de la tectónica de placas supuso una nueva revolución teórica a la que la disciplina de la Paleontología tuvo que adaptarse, abandonando las propuestas de puentes terrestres e hundimientos continentales como maneras de explicar la distribución geográfica de los organismos actuales y pasados. Por otro lado, en el campo de la Biología se produjo un progresivo incremento del interés por los estudios ecológicos, en relación con la creciente percepción del impacto antrópico en los ecosistemas. Dentro de nuestra disciplina, el desarrollo de estas áreas de investigación tuvo como resultado un aumento de los estudios paleobiogeográficos y paleoecológicos (Valentine, 1990). Además, también comenzó por aquel entonces el estudio de las primeras formas de vida terrestre (Awramik, 1971) y se confirmó la preeminencia de la biota de Ediacara (Glaessner, 1959), lo cual propulsó los estudios de fósiles precámbricos. Esas son también las fechas de la reapertura de las excavaciones en Burgess Shale (Briggs et al., 1994; Conway Morris, 1998). Estos trabajos pusieron de manifiesto la complejidad de los eventos evolutivos que dieron lugar a los filos modernos.
 Finalmente, los años 70 del siglo XX vieron el surgir de los estudios macroevolutivos a partir del trabajo de Niles Eldredge y Stephen Jay Gould (1972) sobre el “equilibrio puntuado”, que marcará un cambio de paradigma frente al gradualismo filético imperante hasta el momento. El desarrollo de esta pugna entre gradualismo y puntuacionismo a lo largo de las siguientes décadas ha tenido su reflejo en un debate paralelo sobre las causas últimas de los procesos evolutivos, que pueden verse tanto en las interrelaciones ecológicas dentro de las comunidades, del modo propuesto por ejemplo en la “Teoría de la Reina Roja” de Leigh Van Valen (1973), como en la existencia de factores externos (cambios climáticos, movimientos tectónicos, etc…) que fuerzan la evolución de los organismos como propone Elisabeth S. Vrba (1980, 1992) en la "Teoría del Hábitat". Estos dos enfoques, son herederos de las diferentes perspectivas que mantenían Darwin y Wallace acerca del principal motor evolutivo; mientras el primero enfatizaba la competencia entre individuos para incrementar la supervivencia y la reproducción, el segundo daba mayor preponderancia a la biogeografía y las presiones ambientales sobre las especies. Derivaciones de estas líneas de investigación se pueden ver en los trabajos relacionados con lo que se conoce como “selección de especies” (Stanley, 1975), o el estudio de la diversificación biológica (Sepkoski, 1978) y las extinciones en masa (Raup & Sepkoski, 1984; Jablonski, 1986).
Finalmente, los años 70 del siglo XX vieron el surgir de los estudios macroevolutivos a partir del trabajo de Niles Eldredge y Stephen Jay Gould (1972) sobre el “equilibrio puntuado”, que marcará un cambio de paradigma frente al gradualismo filético imperante hasta el momento. El desarrollo de esta pugna entre gradualismo y puntuacionismo a lo largo de las siguientes décadas ha tenido su reflejo en un debate paralelo sobre las causas últimas de los procesos evolutivos, que pueden verse tanto en las interrelaciones ecológicas dentro de las comunidades, del modo propuesto por ejemplo en la “Teoría de la Reina Roja” de Leigh Van Valen (1973), como en la existencia de factores externos (cambios climáticos, movimientos tectónicos, etc…) que fuerzan la evolución de los organismos como propone Elisabeth S. Vrba (1980, 1992) en la "Teoría del Hábitat". Estos dos enfoques, son herederos de las diferentes perspectivas que mantenían Darwin y Wallace acerca del principal motor evolutivo; mientras el primero enfatizaba la competencia entre individuos para incrementar la supervivencia y la reproducción, el segundo daba mayor preponderancia a la biogeografía y las presiones ambientales sobre las especies. Derivaciones de estas líneas de investigación se pueden ver en los trabajos relacionados con lo que se conoce como “selección de especies” (Stanley, 1975), o el estudio de la diversificación biológica (Sepkoski, 1978) y las extinciones en masa (Raup & Sepkoski, 1984; Jablonski, 1986). En las últimas décadas del siglo XX y en lo que llevamos de siglo XXI el énfasis se ha desplazado hacia el desarrollo de nuevas técnicas y métodos para responder al mismo tipo de preguntas que ya se vienen haciendo desde hace tiempo: ¿Cómo eran y cómo vivían los organismos extintos a los que pertenecen los fósiles que estudiamos? ¿Cómo se relacionan filogenéticamente, entre ellos y con los organismos actuales? ¿Cuál fue la estructura biogeográfica y ecológica de la Biosfera en el pasado? ¿Cómo operan los procesos evolutivos? Para responder a estas cuestiones se está profundizando en aspectos tales como la morfología funcional, la ecomorfología, la filogenia cladista o la modelización cuantitativa y cualitativa, además de establecerse nuevos lazos de colaboración con otras disciplinas con el desarrollo de la microestratigrafía, la geoquímica, la paleosinecología o la morfometría geométrica (Hoffman, 1990).
En las últimas décadas del siglo XX y en lo que llevamos de siglo XXI el énfasis se ha desplazado hacia el desarrollo de nuevas técnicas y métodos para responder al mismo tipo de preguntas que ya se vienen haciendo desde hace tiempo: ¿Cómo eran y cómo vivían los organismos extintos a los que pertenecen los fósiles que estudiamos? ¿Cómo se relacionan filogenéticamente, entre ellos y con los organismos actuales? ¿Cuál fue la estructura biogeográfica y ecológica de la Biosfera en el pasado? ¿Cómo operan los procesos evolutivos? Para responder a estas cuestiones se está profundizando en aspectos tales como la morfología funcional, la ecomorfología, la filogenia cladista o la modelización cuantitativa y cualitativa, además de establecerse nuevos lazos de colaboración con otras disciplinas con el desarrollo de la microestratigrafía, la geoquímica, la paleosinecología o la morfometría geométrica (Hoffman, 1990). 
Referencias
- Awramik, S.M. (1971) Precambrian Columnar Stromatolite Diversity: Reflection of Metazoan Appearance. Science, 174: 825-827.
- Bowler, P.J. (1990) Darwin to Plate Tectonics. En: Paleobiology (eds.: Briggs, D.E.G. & Crowther, P.R.), pp. 543-547. Blackwell, Oxford.
- Briggs, D.E.G., Erwin, D.H. & Collier, F.J. (1994) The Fossils of the Burgess Shale. Smithsonian Institution, Washington.
- Conway Morris, S. (1998) The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford University Press, Oxford.
- Eldredge, N. & Gould, S.J. (1972). Punctuated equilibria: an alternative to phyletic gradualism. En: Models in Paleobiology (ed.: Schopf, T.J.M.), pp. 82-115. Freeman Cooper, San Francisco.
- Glaessner, M.F. (1959) The oldest fossil faunas of South Australia. International Journal of Earth Sciences, 47: 522-531.
- Hoffman, A. (1990) The Past Decade and the Future. En: Paleobiology (eds.: Briggs, D.E.G. & Crowther, P.R.), pp. 550-555. Blackwell, Oxford.
- Jablonski, D. (1986) Background and mass extinctions: the alternation of macroevolutionary regimes. Science, 231: 129-133.
- Raup, D.M. & Sepkoski, J.J. (1984) Periodicity of extinctions in the geologic past. Proceedings of the Nacional Academy of Sciences of the USA, 81: 801-805.
- Sepkoski, J.J. (1978) A kinetic model of Phanerozoic taxonomic diversity. I. Análisis of marine orders. Paleobiology, 4: 223-251.
- Simpson, G.G. (1944) Tempo and Mode in Evolution. Columbia University Press, New York.
- Stanley, S.M. (1975) A theory of evolution above the species level. Proceedings of the Nacional Academy of Sciences of the USA, 72: 646-650.
- Valentine, J.W. (1990) Plate Tectonics to Paleobiology. En: Paleobiology (eds.: Briggs, D.E.G. & Crowther, P.R.), pp. 547-550. Blackwell, Oxford.
- Van Valen, L. (1973) A new evolutionary law. Evolutionary Theory, 1: 1-30.
- Vrba, E.S. (1980) Evolution, species, and fossils: How does life evolve? South African Journal of Science, 76: 61-84.
- Vrba, E.S. (1992) Mammals as a key to evolutionary theory. Journal of Mammalogy, 73: 1-15.


















No hay comentarios:
Publicar un comentario